Cellular Disruption by Ultra-Processed Foods: Mechanistic Pathways Linking Industrial Additives to Metabolic Disease
-
David Chinonso Anih


Department of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba, Nigeria
Verwiyeh Silas TatahDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba, Nigeria
Kenneth Chinekwu UgwuokeDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba, Nigeria
Richard-Harris Nsenreuti BoyiDepartment of Biochemistry, Faculty of Biosciences, Federal University Wukari, Taraba, Nigeria
| Received 29 Aug, 2025 |
Accepted 01 Oct, 2025 |
Published 10 Oct, 2025 |
Ultra-Processed Foods (UPFs) have become a dominant feature of modern diets, coinciding with a global rise in metabolic disorders such as obesity, type 2 diabetes, and cardiovascular disease. While traditionally viewed through the lens of poor nutritional content, emerging research suggests that the health risks of UPFs extend far beyond their macronutrient profiles. This review explores the cellular and molecular mechanisms by which industrial processing and food additives contribute to metabolic dysfunction. It synthesizes evidence from epidemiological studies, clinical trials, and experimental models to present a comprehensive framework linking UPF consumption to systemic and cellular pathology. Central to this framework is the disruption of gut microbiota and intestinal barrier integrity. Additives such as emulsifiers and artificial sweeteners alter microbial composition, reduce beneficial metabolites, and increase gut permeability. These changes facilitate the translocation of endotoxins like lipopolysaccharide into circulation, triggering chronic low-grade inflammation, a key driver of insulin resistance and metabolic disease. Concurrently, preservatives and sweeteners induce oxidative stress, compromising mitochondrial function and cellular energy balance. The cumulative impact of these additives impairs lipid metabolism, glucose homeostasis, and nutrient-sensing pathways, including mTOR, AMPK, and SIRT1, pushing cells toward a state of metabolic overload and dysfunction. This mechanistic insight challenges the conventional nutrient-centric approach to dietary guidelines and underscores the need to consider the nature and extent of food processing. By shifting focus to the biological consequences of industrial formulation, this review advocates for a more holistic understanding of diet-related disease. It calls for public health policies that incorporate food processing classifications, reassess additive safety, and promote minimally processed dietary patterns. Ultimately, the manuscript positions UPFs not merely as unhealthy choices but as active agents of cellular disruption, offering a compelling argument for rethinking how we define and regulate the modern food supply.
| Copyright © 2025 Anih et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
The modern food environment has undergone a profound transformation over the past several decades, characterized by the increasing dominance of Ultra-Processed Foods (UPFs) in diets worldwide. These industrial formulations, which include products such as sweetened beverages, packaged snacks, reconstituted meat products, and pre-prepared frozen meals, are engineered for convenience, hyper-palatability, and long shelf-life. Their proliferation has coincided with a dramatic and concerning increase in the global burden of Non-Communicable Diseases (NCDs), particularly metabolic disorders like obesity, Type 2 Diabetes Mellitus (T2DM), and metabolic syndrome. While the association between UPF consumption and poor health outcomes is well-documented epidemiologically, the underlying biological mechanisms remain a subject of intensive investigation.
Traditionally, the negative health effects of UPFs have been attributed to their poor nutritional profiles-typically high in added sugars, unhealthy fats, and sodium, while being low in dietary fiber, vitamins, and minerals. This nutrient-centric view, however, is increasingly considered incomplete. A growing body of research suggests that the detrimental effects of these foods may be attributable to factors that extend beyond their declared nutrient content1. The very nature of “ultra-processing”, the combination of industrial processes, the disruption of the whole food matrix, and the inclusion of a wide array of cosmetic and functional additives appears to exert unique and potent biological effects. These products are not merely foods with suboptimal nutrient ratios; they are complex industrial formulations whose impact on human physiology must be examined through a different lens2.
Understanding this impact requires a shift in focus from whole-body physiological outcomes to the fundamental cellular and molecular events that precede them. The cell is the primary site where nutrients and non-nutritive compounds are metabolized, and it is at this level that the initial dysfunctions leading to systemic disease originate. Chronic exposure to the unique chemical and physical properties of UPFs has the potential to disrupt cellular homeostasis, impair energy metabolism, and trigger pathological signaling cascades3. These cellular insults, which can include mitochondrial dysfunction, oxidative stress, endoplasmic reticulum (ER) stress, and chronic inflammatory signaling, represent the mechanistic bedrock connecting the consumption of UPFs to the development of metabolic disease. High intake of UPFs has been directly associated with disruptions in metabolism and even cellular degeneration4.
This review seeks to synthesize the current state of knowledge on the cellular metabolic consequences of UPF consumption. Its central thesis is that the combination of industrial processing and the cocktail of additives characteristic of UPFs acts synergistically to disrupt cellular function, initiating a cascade of events that culminates in systemic metabolic dysregulation. To build this argument, this paper will first provide a comprehensive review of the literature defining UPFs and establishing their epidemiological link to chronic disease. It will then explore the literature on two key mediating pathways: The disruption of the gut microbiome and the induction of low-grade systemic inflammation. Following this, the review will delve into the direct cellular impacts of specific food additives and analyze how UPF consumption perturbs key biochemical and signaling pathways, including lipid metabolism, glucose homeostasis, and cellular nutrient sensing. By integrating evidence from epidemiological, clinical, and basic science research, this paper aims to provide a mechanistic framework for understanding how ultra-processed foods contribute to cellular pathology and the modern pandemic of metabolic disease.
Health implications of ultra-processed foods
A review of the literature: The scientific discourse surrounding the health impacts of modern diets has increasingly converged on the concept of food processing. While processing has been part of the human food supply for millennia, the nature, extent, and purpose of contemporary industrial food processing have created a novel category of products with distinct health implications. Before dissecting the specific cellular mechanisms, it is essential to establish a clear definition of UPFs and to review the robust body of evidence linking their consumption to metabolic disease. This section will outline the NOVA classification system as a critical tool for this analysis, summarize the key epidemiological findings, and introduce the gut microbiome and systemic inflammation as primary mediating pathways.
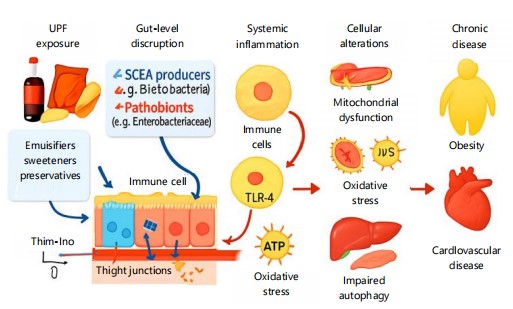
|
Figure 1 illustrates the sequential biological disruptions triggered by Ultra-Processed Food (UPF) exposure, as detailed in Section 2. It begins with gut-level dysbiosis and barrier damage, leading to systemic inflammation and cellular stress responses.
These molecular events culminate in insulin resistance, lipid accumulation, and the onset of chronic diseases. The diagram visually integrates dietary triggers with mechanistic pathways and disease outcomes.
Defining ultra-processed foods
NOVA classification system: A fundamental challenge in studying the health effects of processed foods has been the lack of a standardized and conceptually robust classification system. To address this, a team of researchers at the University of São Paulo developed the NOVA classification system, which categorizes foods not by their nutrient content but “according to the nature, extent, and purpose of the industrial processes they undergo”5. This framework has become the most widely accepted and utilized tool in research on UPFs and has been pivotal in shifting the paradigm from a nutrient-based to a processing-based assessment of food quality. The NOVA system divides all foods and food products into four distinct groups.
Group 1 consists of unprocessed or minimally processed foods. Unprocessed foods are the edible parts of plants or animals in their natural state (e.g., fresh fruits, vegetables, eggs, milk). Minimally processed foods are natural foods that have been altered without the addition of other substances, through processes such as cleaning, grinding, refrigeration, pasteurization, or fermentation. These processes are primarily aimed at preserving the food and making it edible or more accessible.
Group 2 comprises processed culinary ingredients. These are substances derived from Group 1 foods or from nature through processes such as pressing, refining, or grinding. Examples include table sugar from sugarcane, oils from plants, and salt from mines. They are not meant to be consumed on their own but are typically used in combination with Group 1 foods to prepare meals.
Group 3 contains processed foods. These are relatively simple products made by adding Group 2 ingredients (such as salt, sugar, or oil) to Group 1 foods. The main purpose of the processing is to increase the durability of the food or to modify its sensory qualities. Examples include canned fish in oil, fruits in syrup, and freshly made, unpackaged breads and cheeses. While they contain additives, their composition is still recognizable as a modified version of whole foods.
Group 4 is the category of Ultra-Processed Foods (UPFs). This group represents the core of the issue regarding modern dietary patterns. The UPFs are not modified foods but industrial formulations made mostly or entirely from substances extracted from foods (e.g., fats, starches, protein isolates) or synthesized in laboratories (e.g., flavor enhancers, food colorants, emulsifiers). These products typically contain few, if any, whole foods from Group 15. The processes involved are extensive and include extrusion, molding, and pre-frying. The overarching purpose of this ultra-processing is to create highly profitable products (due to cheap ingredients and long shelf-life), convenient (ready-to-eat or ready-to-heat), and hyper-palatable, often engineered to promote consumption. Examples are ubiquitous in the modern food supply and include soft drinks, sweet or savory packaged snacks, instant noodles, reconstituted meat products, and many mass-produced packaged breads and desserts. The use of the NOVA classification allows researchers to isolate the health effects of this specific group of industrial formulations, distinct from other forms of food processing6.
Table 1 presents a comparative overview of the NOVA classification system, which categorizes foods based on processing level and purpose. It outlines definitions, examples, and research relevance for each group, from minimally processed to ultra-processed foods. This framework supports the paradigm shift from nutrient-based to processing-based dietary assessment discussed in Section 2.1.
Epidemiological links to chronic metabolic diseases: Using the NOVA classification framework, a vast and growing body of epidemiological research has uncovered strong and consistent associations between high consumption of UPFs and an increased risk of numerous chronic diseases. Large-scale prospective cohort studies conducted across diverse populations in Europe, North and South America, and Asia have repeatedly demonstrated a dose-response relationship: The higher the proportion of UPFs in the diet, the greater the risk of adverse health outcomes.
One of the most frequently reported associations is with overweight, obesity, and related cardiometabolic conditions. Multiple studies have shown that individuals with higher UPF intake have a greater risk of developing general and abdominal obesity. This link is thought to be mediated not only by the high energy density and poor nutritional quality of UPFs but also by their physical and chemical structure. The soft texture and low fiber content of many UPFs lead to a faster eating rate and diminished satiety signals, promoting passive overconsumption and a greater overall energy intake compared to meals made from unprocessed foods7. This effect on weight gain appears to extend beyond simple caloric content, pointing toward deeper metabolic disturbances7.
Beyond obesity, high UPF consumption has been robustly linked to a higher incidence of type 2 diabetes, cardiovascular diseases, and metabolic syndrome8. These associations often remain significant even after statistical adjustment for established dietary risk factors like saturated fat, sugar, sodium, and dietary fiber intake. This crucial observation suggests that components inherent to the ultra-processing itself, such as the loss of the natural food matrix, the presence of industrial additives, or the formation of neo-formed contaminants during processing, are contributing directly to metabolic pathology. These findings from
population-level studies provide the impetus for investigating the specific biological mechanisms that can explain these powerful and consistent associations. The evidence strongly indicates that UPFs are not passive carriers of “bad nutrients” but may be active promoters of metabolic disease through distinct physiological pathways8.
Table 2 summarizes key prospective cohort studies examining the relationship between Ultra-Processed Food (UPF) intake and chronic metabolic diseases. It highlights study populations, follow-up durations, dietary assessment methods, and reported outcomes such as obesity, type 2 diabetes, and cardiovascular disease. Effect estimates (RR or OR) quantify the strength of these associations. This evidence supports the epidemiological links discussed in Section 2.2.
Gut microbiome as a primary target of UPFs: The human gut is home to a complex ecosystem of trillions of microorganisms, collectively known as the gut microbiome, which plays a fundamental role in host health, including the regulation of nutrient metabolism and immune function9. A healthy and diverse microbiome ferments indigestible dietary fibers to produce beneficial metabolites, most notably Short-Chain Fatty Acids (SCFAs) like butyrate, propionate, and acetate. These SCFAs serve as a primary energy source for intestinal epithelial cells, reinforce the gut barrier, and exert anti-inflammatory effects throughout the body. An increasing body of evidence now points to the gut microbiome as a key mediator of the detrimental health effects of UPFs9.
The composition of UPFs is particularly damaging to a healthy microbiome. They are typically deficient in dietary fiber, the main substrate for beneficial gut bacteria, while being rich in components that can negatively alter the microbial community. The high content of refined sugars and artificial sweeteners can selectively promote the growth of pro-inflammatory bacterial species while reducing overall microbial diversity. Furthermore, many food additives commonly found in UPFs, such as certain emulsifiers and preservatives, have been shown to directly disrupt gut microbial composition and function. This shift in the microbiome, often referred to as dysbiosis, is characterized by a loss of beneficial microbes and a proliferation of potentially pathogenic ones9.
Table 3 presents experimental and observational evidence showing how Ultra-Processed Food (UPF) exposure alters gut microbiome composition and function. It includes both human and animal models, detailing microbial shifts such as reduced SCFA producers and increased pathobionts. These changes are linked to metabolite disruptions like elevated LPS and diminished SCFA levels. The table supports Section 2.3's emphasis on microbiome dysbiosis as a key mediator of UPF-induced metabolic harm.
This dysbiosis has profound consequences for host metabolism. The reduction in fiber-fermenting bacteria leads to decreased production of protective SCFAs, starving the colonic epithelium and impairing its barrier function. Concurrently, the altered microbiome may produce a different profile of metabolites, some of which can be harmful. The disruption of the gut microbiota and its associated metabolism is now considered a likely mechanism through which UPFs impact systemic health10. This microbially-mediated pathway represents a critical link, transforming a dietary exposure into a systemic metabolic and inflammatory challenge, beginning with the integrity of the gut barrier itself10.
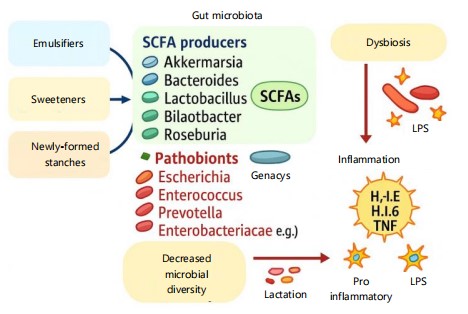
Figure 2 illustrates how specific components of Ultra-Processed Foods (UPFs); emulsifiers, sweeteners, and newly formed starches alter gut microbiota composition. These changes reduce SCFA-producing bacteria and increase pathobionts, leading to decreased microbial diversity. The resulting imbalance promotes inflammation via elevated LPS and pro-inflammatory cytokines. This mechanistic pathway supports the microbial dysbiosis framework discussed in Section 2.3.
| Table 1: | NOVA classification: Food groups, definitions, examples, and research relevance | |||
| NOVA group | Definition | Examples | Relevance to health research | Citation |
| Group 1: MinProc | Edible parts of plants or animals with minimal alteration |
Fresh fruits, vegetables, grains, and meat |
Baseline for healthy dietary patterns; low risk of chronic disease |
Tompa et al.5 and Slaathaug et al.6 |
| Group 2: Culinary ingredients | Substances extracted from group 1 foods or nature for cooking |
Oils, butter, sugar and salt | Used in moderation; not typically consumed alone |
Tompa et al.5 and Slaathaug et al.6 |
| Group 3: Processed foods | Products made by adding group 2 ingredients to group 1 foods |
Canned vegetables, cheese and smoked meats |
Moderate health impact; preservation and flavor enhancement |
Tompa et al.5 and Slaathaug et al.6 |
| Group 4: UPF | Industrial formulations with little to no whole food, often with additives |
Soft drinks, packaged snacks and instant noodles |
Associated with obesity, metabolic syndrome, and other chronic diseases |
Tompa et al.5 and Slaathaug et al.6 |
| MinProc: Minimally processed foods | ||||
| Table 2 | Prospective cohort studies linking UPF intake to metabolic outcomes | |||
| Study | Population | Follow-up duration | UPF measurement method | Outcomes | Effect estimate (RR/OR, CI) | Citation |
| NutriNet-Santé (France) | Adults (n>100,000) | ~5 years | UPF (%) in total diet (24h recall) | Obesity, T2D, CVD | RR: 1.12 (95% CI: 1.06-1.18) | Lv et al.7 and Kityo et al.8 |
| SUN (Spain) | University graduates (n~20,000) | ~10 years | Food frequency questionnaire | Metabolic syndrome, T2D | OR: 1.31 (95% CI: 1.09-1.58) | Lv et al.7 and Kityo et al.8 |
| ELSA-Brazil | Civil servants (n~15,000) | ~8 years | NOVA-based dietary assessment | Hypertension, dyslipidemia | RR: 1.18 (95% CI: 1.05-1.32) | Lv et al.7 and Kityo et al.8 |
| UK Biobank | Adults (n>100,000) | ~10 years | UPF (%) by NOVA classification | All-cause mortality, CVD | HR: 1.24 (95% CI: 1.15-1.34) | Lv et al.7 and Kityo et al.8 |
| RR: Relative risk, OR: Odds ratio and CI: Confidence interval | ||||||
| Table 3 | Human and animal studies reporting microbiome changes with UPF exposure | |||
| Model | Type of UPF exposure | Main microbial shifts | Functional/metabolite outcomes | Citation |
| Human | High UPF diet (>30% energy intake) | ↓ Diversity, ↓ SCFA producers, ↑ pathobionts | ↓ SCFA, ↑ LPS | Rondinella et al.9 and Atzeni et al.10 |
| Animal (mouse) | Emulsifier-enriched chow | ↑ Proteobacteria, ↓ Firmicutes | ↑ LPS, gut permeability | Rondinella et al.9 and Atzeni et al.10 |
| Human | Frequent consumption of sweetened beverages | ↓ Bifidobacteria, ↑ Enterobacteriaceae | ↓ SCFA, ↑ inflammatory markers | Rondinella et al.9 and Atzeni et al.10 |
| Animal (rat) | Processed starch diet | ↓ Lactobacillus, ↑ Clostridium | ↓ SCFA, ↑ oxidative stress | Rondinella et al.9 and Atzeni et al.10 |
| This table summarizes experimental and observational studies examining gut microbiome alterations following UPF exposure. It includes model type, exposure details, microbial shifts, metabolite outcomes, and citations and LPS: Lipopolysaccharide | ||||
|
Induction of systemic low-grade inflammation: Chronic, low-grade inflammation is a well-established underlying driver of metabolic diseases, including insulin resistance, T2DM, and atherosclerosis. This type of inflammation is not a response to acute infection or injury but rather a persistent, low-level activation of the immune system, often triggered by metabolic and nutrient excess11. The consumption of UPFs appears to be a potent trigger for this pathological inflammatory state, acting through several interconnected mechanisms rooted in the gut.
The primary pathway linking UPFs to systemic inflammation is through the disruption of the gut barrier, a condition often termed “leaky gut”. As discussed previously, UPF-induced gut dysbiosis and the direct effects of certain additives can compromise the integrity of the intestinal epithelium. Specifically, these factors can weaken the tight junctions, which are protein complexes that seal the space between epithelial cells and regulate the passage of substances from the gut lumen into the bloodstream. When this barrier is compromised, inflammatory microbial components, such as Lipopolysaccharide (LPS), a component of the outer membrane of Gram-negative bacteria, can translocate from the gut into the circulation. This condition is known as metabolic endotoxemia11.
Once in the bloodstream, LPS is a powerful activator of the innate immune system, binding to receptors like Toll-Like Receptor 4 (TLR4) on immune cells such as macrophages. This binding initiates a signaling cascade that results in the production of pro-inflammatory cytokines, including Tumor Necrosis Factor-Alpha (TNF-α) and Interleukin-6 (IL-6). When this occurs chronically, it establishes a state of systemic low-grade inflammation. This chronic inflammatory environment directly interferes with metabolic processes; for instance, pro-inflammatory cytokines can impair insulin signaling in key metabolic tissues like the liver, muscle, and adipose tissue, leading to insulin resistance. Research has explicitly linked UPF consumption with chronic metabolic inflammation12. This inflammatory state can further alter lipid metabolism and contribute to the cellular degeneration seen in metabolic complications, making it a critical bridge between diet, gut health, and systemic pathology12.
|
| Table 4: | Biomarkers of inflammation associated with UPF consumption | |||
| Biomarker | Reported change | Sample type | Study model | Citation |
| CRP | ↑ Concentration | Blood serum | Human cohort | Kotopoulou et al.11 and Martins-Gomes et al.12 |
| IL-6 | ↑ Expression | Plasma | Animal model | Kotopoulou et al.11 and Martins-Gomes et al.12 |
| TNF-α | ↑ Circulating levels | Blood serum | Human cohort | Kotopoulou et al.11 and Martins-Gomes et al.12 |
| IL-6, TNF-α | ↑ Gene transcription | Intestinal tissue | Animal model | Kotopoulou et al.11 and Martins-Gomes et al.12 |
| This table presents inflammatory markers commonly elevated in response to UPF exposure. It includes reported changes, sample types, study models, and citations. CRP: C-Reactive Protein, IL-6: Interleukin-6 and TNF-α: Tumor Necrosis Factor-Alpha | ||||
Table 4 highlights inflammatory biomarkers elevated in studies examining UPF consumption, supporting the mechanistic link to systemic low-grade inflammation discussed in Section 2.4. It includes both human and animal models, with sample types ranging from blood serum to intestinal tissue. The consistent rise in CRP, IL-6, and TNF-α underscores UPFs’ role in triggering immune activation. Citations anchor each finding in peer-reviewed research.
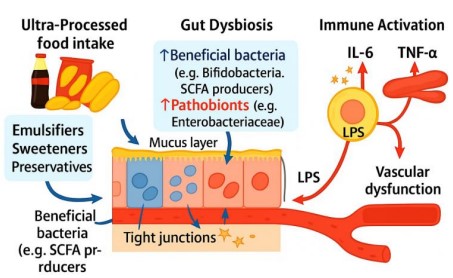
Figure 3 illustrates the pathway described in Section 3.3, showing how Ultra-Processed Food (UPF) consumption disrupts gut microbiota balance.
This dysbiosis compromises intestinal barrier integrity, enabling translocation of Lipopolysaccharides (LPS) into circulation.
The LPS triggers immune activation via TLR4, elevating pro-inflammatory cytokines (IL-6, TNF-α), which contribute to insulin resistance and vascular dysfunction.
The diagram visually integrates microbial shifts, epithelial damage, and systemic inflammatory signaling.
Impact of food additives on cellular homeostasis: Ultra-Processed Foods (UPFs) are distinct not only due to their nutrient profiles but fundamentally because they are industrial formulations containing a host of non-culinary substances known as food additives. These additives, including emulsifiers, preservatives, artificial sweeteners, stabilizers, and colorants, are incorporated to enhance palatability, extend shelf life, and improve texture. While generally recognized as safe at approved concentrations, a growing body of evidence suggests that chronic, cumulative exposure to these compounds can have profound and deleterious effects on cellular homeostasis. Food additives are no longer viewed as inert passengers in the food matrix; instead, they are increasingly identified as bioactive agents capable of directly interacting with and perturbing fundamental biological processes. Their impact begins at the first point of contact, the gastrointestinal tract, and extends to systemic effects on cellular energy production, redox balance, and inflammatory signaling. Understanding the cellular-level effects of these additives is critical to unravelling the mechanisms by which UPFs contribute to metabolic disease13. This section will dissect the specific impacts of key classes of food additives on gut barrier integrity, oxidative stress, and mitochondrial function, revealing them as significant mediators of UPF-induced cellular pathology.
Emulsifiers, gut barrier integrity, and metabolic endotoxemia: Emulsifiers are ubiquitous in UPFs, added to improve texture and prevent the separation of oil and water components in products like baked goods, ice cream, and salad dressings. These amphipathic molecules, which include synthetic compounds such as Carboxymethylcellulose (CMC) and polysorbate 80 (P80), possess detergent-like properties that are central to their technological function and, consequently, their biological impact. The intestinal epithelium is protected by a multi-layered mucus barrier that segregates the vast luminal microbial community from the host’s underlying tissues. Emerging research indicates that dietary emulsifiers can directly disrupt this critical defensive layer. By virtue of their surface-active nature, emulsifiers can erode the stratified mucus gel, reducing its thickness and integrity, thereby allowing bacteria to encroach upon the epithelial cell surface, a region that is normally kept sterile14.
This physical degradation of the mucus barrier is often coupled with a direct alteration of the gut microbiota’s composition and function. Studies have shown that commonly used emulsifiers can selectively promote the growth of pro-inflammatory, mucus-degrading bacterial species while diminishing populations of beneficial microbes14. For instance, exposure to CMC and P80 has been associated with an increased abundance of bacteria capable of penetrating the mucus layer and expressing pro-inflammatory molecules14. This dysbiotic shift in the microbial ecosystem further contributes to the breakdown of the intestinal barrier.
Beyond their effects on mucus and microbes, emulsifiers appear to directly impact the integrity of the epithelial cell layer itself. The intestinal epithelium forms a selective barrier through a series of intercellular protein complexes known as tight junctions, which regulate paracellular transport. Research has demonstrated that various emulsifiers can compromise the function of these junctions, leading to increased intestinal permeability14,15. Some studies have shown that emulsifiers can increase bacterial adhesion to human intestinal epithelial cell monolayers, suggesting a direct interaction that could facilitate pathogen translocation or inflammatory responses14,15. The consequence of this compromised barrier function is a condition known as “leaky gut”, where components from the gut lumen that are normally contained, such as microbial fragments, can pass into the bloodstream.
Of particular importance is the translocation of Lipopolysaccharide (LPS), a major component of the outer membrane of Gram-negative bacteria. When LPS enters systemic circulation, it triggers a potent inflammatory response, a state termed metabolic endotoxemia. This condition is characterized by chronically elevated, low-grade levels of circulating endotoxins that activate the innate immune system, leading to the production of pro-inflammatory cytokines like TNF-α and IL-615. Metabolic endotoxemia is now recognized as a key mechanistic link between gut-derived insults and the development of systemic metabolic diseases, including insulin resistance, hepatic steatosis, and chronic inflammation. Therefore, by disrupting gut microbiota balance and impairing gut barrier integrity, emulsifiers and other additives found in UPFs directly contribute to a pro-inflammatory systemic environment that drives cellular metabolic dysfunction across multiple organ systems15.
Preservatives and sweeteners
Inducers of oxidative stress: Beyond emulsifiers, UPFs are formulated with a wide array of preservatives and synthetic sweeteners designed to enhance shelf stability and palatability while reducing caloric content. Preservatives, such as sulfites, benzoates, and sorbates, function by creating an environment hostile to microbial growth, while non-nutritive sweeteners provide intense sweetness without the energy of sugar. However, the biochemical properties that make these substances effective can also impose a significant burden on cellular redox homeostasis. Growing evidence indicates that many of these common additives can act as potent inducers of oxidative stress, a state of imbalance between the production of reactive oxygen species (ROS) and the cell’s capacity to neutralize them through antioxidant defenses16.
Oxidative stress represents a fundamental mechanism of cellular damage. The ROS, which include superoxide anions, hydrogen peroxide, and hydroxyl radicals, are highly reactive molecules that can indiscriminately damage essential cellular macromolecules. When produced in excess, they can trigger lipid peroxidation, a process that degrades cellular membranes, compromises their fluidity and function, and generates further toxic byproducts. Proteins can undergo carbonylation and other oxidative modifications, leading to conformational changes, loss of enzymatic function, and aggregation. The DNA is also vulnerable, with oxidative damage to nucleic acids causing mutations that can impair cellular function and replication. A number of food additives have been implicated in promoting this damaging state16.
Synthetic preservatives, for instance, may induce oxidative stress through several mechanisms. Some may directly participate in redox cycling reactions that generate ROS, while others may deplete the cell’s primary endogenous antioxidant, Glutathione (GSH), by forming conjugates with it, thereby weakening the cell’s overall antioxidant capacity. The antimicrobial properties of preservatives, while targeted at bacteria and fungi, may not be entirely specific. These compounds can have off-target effects on host cell mitochondria, which share evolutionary origins with bacteria. By interfering with mitochondrial processes, preservatives can cause electrons to leak from the electron transport chain, directly generating superoxide radicals and initiating a cascade of oxidative damage16,17. This suggests that additives intended for food preservation can inadvertently predispose cells to oxidative stress16,17.
Similarly, artificial sweeteners have been linked to increased oxidative stress, although the mechanisms are still under investigation. Some research suggests that the metabolism of certain sweeteners can generate ROS as byproducts. More significantly, like emulsifiers, sweeteners profoundly alter the composition and metabolic output of the gut microbiome. This dysbiosis can lead to a reduction in the production of beneficial microbial metabolites, such as Short-Chain Fatty Acids (SCFAs) that possess antioxidant properties, and an increase in pro-inflammatory signals that themselves promote oxidative stress systemically17. The chronic consumption of these additives, common in diets high in UPFs, can therefore lead to a sustained state of low-grade oxidative stress throughout the body. This persistent oxidative damage is a key contributor to cellular aging, inflammation, and the initiation and progression of chronic diseases, including metabolic disorders and neurodegenerative conditions17. The combined and potentially synergistic effects of multiple additives within a single food product may create a significant oxidative burden that challenges cellular resilience and contributes directly to cellular pathology17.
Table 5 presents evidence linking specific food additives to oxidative stress, supporting the mechanistic insights discussed in Section 3.2 of the manuscript. It highlights how preservatives and sweeteners contribute to ROS production, antioxidant depletion, and mitochondrial damage. The table includes experimental models and citations to anchor each finding. These mechanisms underscore the cellular risks posed by chronic UPF additive exposure.
| Table 5: | Additives linked to oxidative stress: Mechanisms and supporting evidence | |||
| Additive type | Specific compound | Reported mechanism | Model | Citation |
| Preservative | Sodium benzoate | ↑ ROS, ↓ GSH, mitochondrial membrane disruption |
Animal (Rat) | Dey et al.16 and Jarmakiewicz-Czaja et al.17 |
| Preservative | Potassium sorbate | Lipid peroxidation, ↑ MDA levels | Cell culture | Dey et al.16 and Jarmakiewicz-Czaja et al.17 |
| Sweetener | Aspartame | ↑ ROS, mitochondrial uncoupling | Animal (Mouse) | Dey et al.16 and Jarmakiewicz-Czaja et al.17 |
| Sweetener | Sucralose | ↓ GSH, altered gut microbiota → systemic oxidative stress |
Human (Pilot) | Dey et al.16 and Jarmakiewicz-Czaja et al.17 |
| This table outlines preservatives and sweeteners associated with oxidative stress. It includes mechanisms such as ROS generation, antioxidant depletion, and mitochondrial disruption across various models. ROS: Reactive oxygen species, MDA: Malondialdehyde and GSH: Reduced glutathione | ||||
Additive-induced mitochondrial dysfunction and energy imbalance: Mitochondria are dynamic organelles at the heart of cellular metabolism, responsible for generating over 90% of the cell’s adenosine triphosphate (ATP) through oxidative phosphorylation. They are not only the cell’s powerhouses but also central hubs for signaling, redox balance, and apoptosis. Due to their high metabolic rate and central role in oxygen consumption, mitochondria are both a primary source of endogenous reactive oxygen species (ROS) and a primary target of oxidative damage. The oxidative stress induced by food additives, as discussed previously, therefore has a particularly devastating impact on mitochondrial integrity and function18. A growing body of evidence now indicates that various additives, including preservatives and sweeteners, can directly or indirectly trigger mitochondrial dysfunction, leading to a profound cellular energy imbalance and initiating a vicious cycle of further damage18.
The assault on mitochondria can occur through multiple pathways. First, the increased ROS generated by additives can directly attack mitochondrial components. The inner mitochondrial membrane, rich in polyunsaturated fatty acids, is highly susceptible to lipid peroxidation, which can compromise its integrity and disrupt the electrochemical proton gradient essential for ATP synthesis. Key proteins of the electron transport chain (ETC) and other mitochondrial enzymes can be oxidatively damaged, leading to their inactivation and a subsequent reduction in respiratory efficiency19. Furthermore, Mitochondrial DNA (mtDNA), which lacks the protective histone proteins and robust repair mechanisms of nuclear DNA, is particularly vulnerable to oxidative damage. Mutations in mtDNA can lead to the synthesis of defective ETC components, permanently impairing the organelle’s function.
Beyond indirect damage via oxidative stress, some food additives may exert a more direct toxic effect on mitochondria. Certain compounds may act as inhibitors of specific ETC complexes, physically blocking the flow of electrons and crippling ATP production. Others may function as “uncouplers”, molecules that dissipate the proton gradient across the inner mitochondrial membrane without the concomitant production of ATP. This uncoupling leads to energy being lost as heat and forces the ETC to work harder to maintain the gradient, a process that paradoxically increases ROS generation and oxygen consumption while failing to produce usable energy. This creates a state of severe bioenergetic crisis within the cell, where energy demand outstrips supply19.
Table 6 compiles evidence from human, animal, and cell-based studies showing how UPF additives impair mitochondrial function, as discussed in Section 3.3. Key endpoints include reduced ATP production, inhibited ETC activity, and altered oxygen consumption. These findings highlight mitochondrial vulnerability to additive-induced stress and energy imbalance.
The consequences of additive-induced mitochondrial dysfunction are far-reaching. Impaired ATP production compromises all energy-dependent cellular processes, from muscle contraction and nerve impulse transmission to DNA repair and protein synthesis. To compensate for the failure of oxidative
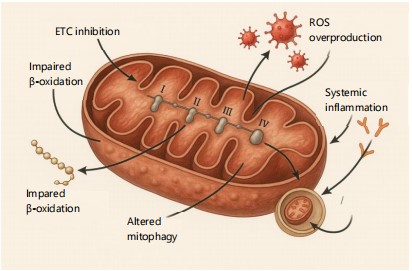
|
Mitochondrion is shown with labeled dysfunction sites caused by UPF additives. Key disruptions include ETC inhibition, excessive ROS generation, impaired fatty acid oxidation, and altered mitophagy. These changes collectively result in reduced ATP output and promote inflammatory signaling. ETC: Electron transport chain and ROS: Reactive oxygen species phosphorylation, cells may shift their metabolism towards less efficient pathways like aerobic glycolysis, a hallmark of many pathological states. Furthermore, damaged mitochondria become a major source of pro-inflammatory and pro-apoptotic signals. They can release Danger-Associated Molecular Patterns (DAMPs), such as fragments of mtDNA and cardiolipin, which activate innate immune responses and perpetuate inflammation. Persistent mitochondrial damage can also trigger the intrinsic pathway of apoptosis, leading to programmed cell death. This creates a self-amplifying cycle: Additive-induced oxidative stress causes mitochondrial dysfunction, which in turn leads to more ROS production, further cellular damage, and ultimately, cell death and tissue degradation19. This cascade positions mitochondrial dysfunction as a critical nexus between food additive exposure and the development of chronic metabolic diseases.
| Table 6: | Evidence of mitochondrial dysfunction linked to UPF additives | |||
| Study model | UPF component | Mitochondrial endpoint affected | Reported effect | Citation |
| Animal (Rat) | Sodium nitrite | ATP levels | ↓ ATP production | Hrncir et al.18 and Claudino et al.19 |
| Cell culture | Aspartame | OCR, membrane potential | ↓ OCR, disrupted membrane integrity | Hrncir et al.18 and Claudino et al.19 |
| Animal (Mouse) | Emulsifiers (CMC, P80) |
ETC activity | ETC inhibition, ↑ ROS generation | Hrncir et al.18 and Claudino et al.19 |
| Human (Pilot) | Mixed sweeteners | ATP levels, OCR | ↓ ATP, altered oxygen consumption | Hrncir et al.18 and Claudino et al.19 |
| This table presents studies examining mitochondrial endpoints affected by UPF components. It includes model type, specific additives, affected parameters, and reported effects. OCR: Oxygen consumption rate and ETC: Electron transport chain | ||||
Figure 4 illustrates the major sites of mitochondrial damage triggered by Ultra-Processed Food (UPF) additives, as discussed in Section 3.3. The mechanisms include electron transport chain (ETC) inhibition, ROS overproduction, impaired β-oxidation, and disrupted mitophagy. These dysfunctions compromise ATP production and generate pro-inflammatory signals. Together, they highlight the central role of mitochondrial decay in UPF-induced energy imbalance.
Key biochemical pathways disrupted by ultra-processed foods: The adverse health effects of Ultra-Processed Foods (UPFs) are not solely attributable to the actions of individual food additives but are also a consequence of their overall composition and the profound impact of industrial processing on the food matrix. The UPFs are typically characterized by high energy density, an abundance of rapidly absorbable free sugars (especially fructose), unhealthy fats, and a low content of fiber and micronutrients. This unique nutritional profile, combined with the metabolic insults from additives, converges to disrupt the body’s most fundamental biochemical pathways. The chronic consumption of these foods overwhelms cellular metabolic capacity, subverts intricate regulatory networks, and promotes a pathological state conducive to disease. This section explores the disruption of three central pillars of cellular metabolism: lipid metabolism and its link to hepatic steatosis, glucose homeostasis and insulin signaling, and the overarching nutrient-sensing pathways that govern cellular growth and energy balance.
Aberrations in lipid metabolism and hepatic steatosis: The liver is the body’s primary metabolic hub, playing a central role in processing dietary nutrients and regulating lipid homeostasis. Chronic consumption of UPFs places an exceptional burden on this organ, directly promoting aberrations in lipid metabolism that culminate in hepatic steatosis, or the accumulation of fat within liver cells (hepatocytes). This condition, also known as Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD), is a foundational step in the progression toward more severe liver diseases like steatohepatitis, fibrosis, and cirrhosis20. The link between UPFs and hepatic steatosis is multifactorial, stemming from both the macronutrient composition of the foods and the bioactivity of their additives.
A key driver of this process is the high content of fructose, often in the form of high-fructose corn syrup, found in many sweetened beverages and processed snacks. Unlike glucose, which can be utilized by most cells in the body and is subject to tight regulation by insulin, fructose is almost exclusively metabolized by the liver. Importantly, its metabolism bypasses the main rate-limiting step of glycolysis, phosphofructokinase, allowing it to provide an unregulated and rapid influx of carbon substrates into hepatic metabolic pathways. This flood of substrates, far exceeding the liver’s immediate energy needs, is preferentially shunted into the pathway of de novo lipogenesis (DNL), the synthesis of new fatty acids from non-lipid precursors. The resulting fatty acids are then esterified into triglycerides, which accumulate within hepatocytes as lipid droplets20.
This process is exacerbated by the state of systemic low-grade inflammation and insulin resistance induced by UPFs. As established, components of UPFs can compromise the gut barrier, leading to metabolic endotoxemia21. The resulting inflammation, mediated by cytokines like TNF-α, directly promotes hepatic insulin resistance. In a paradoxical state of “selective insulin resistance”, the liver becomes resistant to insulin’s suppressive effect on glucose production but remains sensitive to its stimulatory effect on lipogenesis. This means the liver continues to produce glucose, contributing to hyperglycemia, while simultaneously ramping up fat production. This dysregulation is a central feature of metabolic syndrome and underscores how UPFs can simultaneously impair glucose and lipid control21.
Furthermore, the overall metabolic context created by UPF consumption shifts the balance of lipid handling in the liver. Beyond increased DNL, there may be an impairment in the liver’s ability to export triglycerides in the form of Very-Low-Density Lipoprotein (VLDL) and a reduction in fatty acid oxidation (the burning of fat for energy). The suppression of key regulatory molecules like AMPK (as will be discussed later) further tilts the balance away from catabolism and toward lipid storage. This results in an abnormal fatty acid metabolism profile, not just in the liver but systemically, contributing to dyslipidemia characterized by high circulating triglycerides and altered cholesterol profiles22. The cumulative effect is that UPFs program the liver for fat accumulation, creating a cellular environment ripe for further injury, inflammation, and the progression of chronic metabolic disease22.
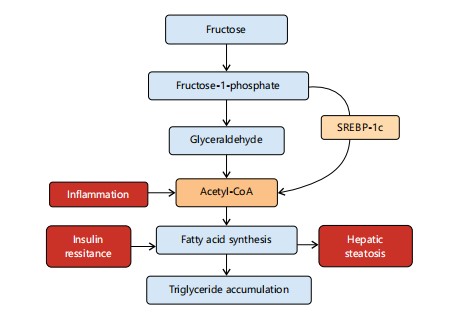
|
Figure 5 shows how excess fructose from ultra-processed foods enters hepatic metabolism, bypassing regulatory control. It highlights the conversion to acetyl-CoA and subsequent stimulation of fatty acid synthesis through SREBP-1c activation. Inflammatory signaling and insulin resistance further amplify lipogenesis. These events culminate in triglyceride accumulation and hepatic steatosis, as discussed in Section 4.1.
Impaired glucose homeostasis and glycolytic pathways: The regulation of blood glucose concentration is a cornerstone of metabolic health, orchestrated primarily by the hormone insulin. A diet high in UPFs systematically undermines this delicate balance, leading to impaired glucose homeostasis and the development of insulin resistance, a precursor to type 2 diabetes. This disruption occurs at multiple levels, from the rapid absorption of refined carbohydrates in the gut to the desensitization of insulin signaling pathways within target cells such as muscle, liver, and adipose tissue23.
The UPFs are typically engineered to have a high glycemic index and glycemic load, meaning they are rapidly digested and absorbed, causing sharp, pronounced spikes in postprandial blood glucose. This hyperglycemia triggers a correspondingly large and rapid release of insulin from the pancreas. Over time, chronic exposure to these exaggerated cycles of hyperglycemia and hyperinsulinemia places immense stress on the metabolic system. Target cells, in an attempt to protect themselves from glucose toxicity and the overstimulation by insulin, begin to downregulate their response to the hormone. This is the essence of insulin resistance23.
At the cellular level, insulin resistance manifests as a failure of the insulin signaling cascade. When insulin binds to its receptor on the cell surface, it initiates a series of phosphorylation events, primarily through the IRS-PI3K-Akt pathway. A key downstream effect of this pathway in muscle and fat cells is the translocation of the Glucose Transporter protein 4 (GLUT4) from intracellular vesicles to the plasma membrane24. The GLUT4 is the primary transporter responsible for insulin-stimulated glucose uptake. In a state of insulin resistance, this signaling pathway is blunted. Inflammatory cytokines (resulting from metabolic endotoxemia) and intracellular lipid accumulation (lipotoxicity) can phosphorylate inhibitory
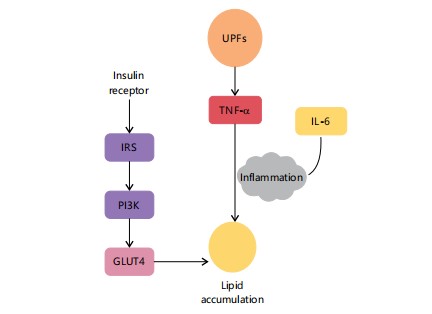
|
Diagram shows the insulin signaling cascade (Insulin Receptor → IRS → PI3K → GLUT4) and its disruption by inflammatory mediators. UPFs activate TNF-α and IL-6, enhancing inflammation and lipid accumulation. IRS: Insulin receptor substrate, PI3K: Phosphoinositide 3-Kinase, GLUT4: Glucose Transporter Type 4, TNF-α: Tumor Necrosis Factor-Alpha and IL-6: Interleukin-6 sites on insulin receptor substrates (IRS), effectively blocking the signal. Consequently, GLUT4 fails to properly translocate to the cell surface, and glucose uptake from the blood is severely impaired. This leaves glucose circulating at high levels, perpetuating the vicious cycle of hyperinsulinemia and worsening insulin resistance24.
Furthermore, there is evidence that UPFs can disrupt glycolytic metabolism itself, the fundamental ten-step pathway that breaks down glucose to produce ATP and metabolic intermediates25. This could involve alterations in the expression or activity of key glycolytic enzymes. The gut microbiome, profoundly altered by UPF additives, also plays a role. Dysbiosis can impact glucose metabolism systemically, and specific food preservatives have been shown to directly interfere with glucose metabolism in experimental models25. The aggregate effect is a profound dysregulation where peripheral tissues are effectively “starved” of glucose in the midst of plenty, while the insulin-resistant liver fails to suppress its own glucose production (gluconeogenesis), further contributing to high blood sugar levels. This cellular-level breakdown in glucose handling is a hallmark of the metabolic disruption caused by UPF consumption.
Figure 6 illustrates how Ultra-Processed Foods (UPFs) impair insulin signaling, contributing to lipid accumulation. It contrasts the normal insulin pathway involving IRS, PI3K, and GLUT4 with inflammatory interference. The TNF-α and IL-6, induced by UPFs, disrupt signaling and promote chronic inflammation. These molecular alterations lead to impaired glucose uptake and metabolic dysfunction as outlined in Section 4.2.
Dysregulation of cellular nutrient sensing and signaling: At the core of cellular metabolic regulation lie intricate nutrient-sensing pathways that act as master switches, interpreting the availability of energy and nutrients to orchestrate an appropriate response, be it growth, maintenance, or survival. These pathways, including mTOR (mammalian target of rapamycin), AMPK (AMP-activated protein kinase), and the sirtuins, form a complex signaling network that maintains metabolic homeostasis. A diet rich in UPFs, characterized by a relentless surplus of energy and specific macronutrients, systematically dysregulates this network, locking cells into a pathological state of constant anabolic signaling that promotes disease26.
The mTOR pathway, specifically the mTORC1 complex, is a central regulator of cell growth, proliferation, and anabolism. It is activated by a convergence of signals, including growth factors (like insulin) and high levels of nutrients, particularly amino acids and glucose. The chronic hyperinsulinemia and nutrient excess associated with UPF consumption lead to the persistent, unabated activation of mTORC1. While essential for growth, chronic mTORC1 activation is detrimental. It potently stimulates anabolic processes like protein synthesis and de novo lipogenesis, contributing to cellular hypertrophy and fat accumulation. Simultaneously, and perhaps more critically, activated mTORC1 is a powerful inhibitor of autophagy26. Autophagy is the cell’s essential housekeeping process, responsible for degrading and recycling damaged organelles (like dysfunctional mitochondria), misfolded proteins, and other cellular debris. By suppressing autophagy, chronic mTORC1 activation allows this cellular “garbage” to accumulate, leading to increased cellular stress, inflammation, and dysfunction26.
In direct opposition to mTOR is the AMPK pathway. AMPK functions as the cell’s primary energy sensor, activated by a high AMP: ATP ratio, which signals a low-energy state. Once activated, AMPK works to restore energy balance by switching on catabolic, ATP-producing pathways (such as fatty acid oxidation and glucose uptake) while switching off anabolic, ATP-consuming pathways (like lipid and protein synthesis)27. The constant energy surplus provided by UPFs leads to a chronically high cellular energy charge (a low AMP: ATP ratio), which robustly suppresses AMPK activity. This suppression effectively removes the brakes on lipid synthesis and turns off the machinery for fat burning, directly contributing to the development of hepatic steatosis and obesity27.
The sirtuins are another critical class of metabolic regulators, a family of NAD+-dependent deacetylases that link cellular energy status to the regulation of gene expression and protein function. Sirtuin 1 (SIRT1), a key member of this family, is activated by a high NAD+/NADH ratio, a condition associated with energy deficit (e.g., during fasting or exercise). Activated SIRT1 orchestrates a broad program of metabolic fitness, promoting mitochondrial biogenesis, enhancing insulin sensitivity, and suppressing inflammation28. The metabolic state induced by UPF consumption, characterized by a constant influx of reducing equivalents from the metabolism of sugars and fats, leads to a low NAD+/NADH ratio, which inhibits SIRT1 activity. The disruption of SIRT1 function has been directly implicated in the dysregulation of metabolic and other cellular functions, contributing to the pathophysiology of chronic disease28. By subverting these three fundamental nutrient-sensing pathways, UPFs reprogram cellular metabolism toward a state of relentless synthesis, storage, and suppressed maintenance, creating the biochemical foundation for the development of a spectrum of metabolic disorders28.
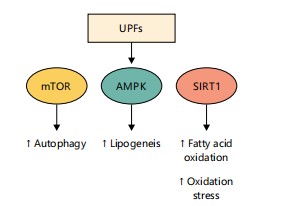
Figure 7 illustrates how Ultra-Processed Foods (UPFs) alter key cellular nutrient-sensing pathways, as detailed in Section 4.3. It shows chronic activation of mTOR, suppression of AMPK regulation, and inhibition of SIRT1 activity under UPF consumption. These disruptions promote lipogenesis, suppress autophagy, reduce fatty acid oxidation, and increase oxidative stress. Such dysregulation drives metabolic imbalance and cellular dysfunction.
From cellular mechanisms to public health action
Integrative discussion and research imperatives on ultra-processed foods: This Section integrates mechanistic insights into a unified cascade from gut barrier disruption through systemic inflammation to cellular energy failure. It highlights how industrial additives, including emulsifiers, sweeteners, and preservatives, drive oxidative stress, mitochondrial dysfunction, and insulin resistance. The discussion reframes dietary advice by shifting from nutrient-centric guidelines to processing-aware frameworks that recognize UPFs as active disease agents. It advocates for policy actions including clear front-of-package labeling, additive safety reevaluation, and fiscal measures to promote minimally processed foods. Key limitations are addressed, such as overreliance on in vitro and animal models and the need to understand additive cocktail effects in human contexts. Future research priorities emphasize long-term clinical trials, advanced organ-on-a-chip models, multi-omics profiling, and development of refined food processing classifications.
|
Synthesis of mechanisms
From processing to cellular pathology: The evidence reviewed herein delineates a complex, multi-layered mechanistic pathway through which Ultra-Processed Foods (UPFs) contribute to metabolic dysregulation and cellular pathology29. The adverse health outcomes associated with UPF consumption are not merely a consequence of their poor macronutrient profiles high in added sugars, unhealthy fats, and sodium, but are fundamentally rooted in the physical and chemical alterations inherent to industrial processing and the functional effects of a wide array of additives. This synthesis moves beyond a nutrient-centric view to a holistic model where the processing itself is a primary determinant of cellular harm29.
The initial interface for this pathogenic cascade is the gastrointestinal tract. Unlike whole or minimally processed foods, the UPF matrix is often designed for hyperpalatability and rapid digestion, leading to accelerated nutrient absorption and subsequent sharp fluctuations in postprandial glucose and insulin. Concurrently, specific additives, particularly emulsifiers, directly challenge the integrity of the intestinal epithelial barrier30. These compounds can alter the composition and function of the gut microbiota and disrupt the protective mucus layer, increasing intestinal permeability. This breach of the gut barrier facilitates the translocation of pro-inflammatory microbial components, such as Lipopolysaccharide (LPS), from the gut lumen into systemic circulation, a condition known as metabolic endotoxemia.
This low-grade systemic inflammation becomes a critical driver of downstream cellular dysfunction. Circulating inflammatory mediators activate signaling cascades, such as the NF-κB pathway, in metabolically active tissues like the liver, adipose tissue, and skeletal muscle. This inflammatory milieu directly contributes to the development of insulin resistance, where cells become less responsive to insulin’s signals to uptake glucose30. Furthermore, the constant immunological challenge, combined with the metabolic burden of processing excessive refined carbohydrates and fats, promotes a state of chronic oxidative stress. Many food additives, including certain preservatives and artificial sweeteners, have been implicated in exacerbating this oxidative burden by either generating reactive oxygen species (ROS) or impairing the cell’s endogenous antioxidant defense systems30.
Table 7 synthesizes mechanistic insights from prior sections, linking UPF exposure to biological disruptions across gut, systemic, and cellular levels. It evaluates the robustness of current evidence and highlights key gaps in experimental and epidemiological research. This mapping supports the integrative framework presented in Section 5.1.
| Table 7: | Mapping of mechanisms, evidence strength, and key research gaps | |||
| Mechanism | Evidence strength | Key research gaps | Citation |
| Microbiome dysbiosis | Strong | Longitudinal human studies; strain-level resolution |
|
| Systemic inflammation | Moderate | Biomarker standardization; causal inference in humans |
|
| Oxidative stress | Moderate | Dose-response data;additive-specific effects | |
| Mitochondrial dysfunction | Weak | Human validation; mechanistic pathways across cell types |
|
| This table outlines major biological mechanisms affected by UPFs, categorizing evidence strength and identifying research gaps. Mechanisms include microbiome dysbiosis, inflammation, oxidative stress, mitochondrial dysfunction and evidence strength is classified as strong, moderate, or weak | |||
At the subcellular level, the mitochondrion emerges as a central target of UPF-induced damage. Mitochondrial integrity is essential for energy homeostasis, and its dysfunction is a hallmark of metabolic disease. Research indicates that various food additives can directly interfere with mitochondrial function, disrupting the electron transport chain, reducing ATP synthesis, and increasing ROS leakage. This energy imbalance, coupled with heightened oxidative stress, creates a vicious cycle that damages mitochondrial DNA, proteins, and lipids, leading to further mitochondrial decay and cellular apoptosis. This process helps explain the profound link between high UPF consumption and outcomes such as cellular degeneration and death. Ultimately, the journey from the industrial formulation of a UPF to the disruption of a hepatocyte’s mitochondrial respiration illustrates a clear pathological trajectory driven by the cumulative effects of processing and additives, which fundamentally alter nutrient metabolism and cellular homeostasis25.
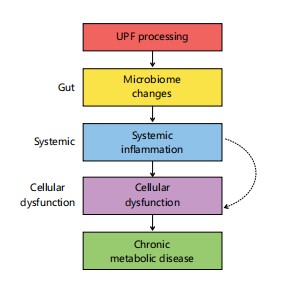
Figure 8 presents a unified mechanistic flowchart tracing the progression from UPF processing to chronic metabolic disease. It integrates gut-level disruptions, systemic inflammation, and cellular dysfunction into a coherent biological cascade. Solid arrows represent established causal pathways, while dashed arrows indicate hypothetical links. This synthesis reflects the multi-layered framework discussed in Section 5.1.
Implications for public health and dietary guidelines: The cellular and molecular mechanisms linking UPFs to metabolic disease carry profound implications for public health policy and dietary guidance. For decades, nutritional advice has been dominated by a reductionist focus on individual nutrients, encouraging the public to limit saturated fat, added sugar, and sodium26. While this advice remains relevant, the evidence on UPFs suggests it is critically insufficient. A food product can be formulated to be low in these “nutrients of concern” yet still be ultra-processed and contain a cocktail of additives that disrupt gut health, promote inflammation, and impair mitochondrial function. Therefore, dietary guidelines that ignore the extent and purpose of food processing fail to address a key driver of diet-related chronic disease.
A necessary evolution in public health nutrition is the integration of food processing classifications, such as the NOVA system, into national dietary recommendations. Informing the public that not all processed foods are equal and providing clear definitions of UPFs can empower consumers to make more informed choices. This requires a paradigm shift from simply reading a nutrition facts panel to also considering the ingredient list and the nature of the product itself. Public health campaigns should be designed to educate citizens about the risks associated with the “ultra-processing” phenomenon, highlighting that the convenience and low cost of these products may come with significant long-term biological costs27.
Beyond consumer education, policy-level interventions are warranted. These could include front-of-package labeling systems that clearly identify UPFs, fiscal policies such as taxes on UPFs or subsidies for fresh and minimally processed foods, and restrictions on the marketing of UPFs, particularly to children.
|
Furthermore, regulatory bodies should re-evaluate the safety of certain food additives based on their metabolic and gut microbiome effects, rather than solely on traditional toxicological endpoints. While some initiatives explore the possibility of “re-engineering” UPFs to be less harmful, for instance by improving their “Metabolic Matrix,” such approaches must be viewed with caution. While potentially beneficial, this focus on technological fixes should not divert attention from the more pressing public health priority: Promoting dietary patterns centered on whole and minimally processed foods and reducing the overall consumption of industrial formulations28.
LIMITATIONS AND FUTURE RESEARCH DIRECTIONS
Despite the compelling evidence linking UPF consumption to adverse metabolic outcomes, the field is characterized by several limitations that must be addressed in future research. A primary challenge lies in translating findings from in vitro and animal models to human pathophysiology. Many mechanistic studies utilize isolated cell lines or rodent models exposed to individual additives at concentrations that may or may not reflect typical human dietary intake. While essential for establishing biological plausibility, these models cannot fully replicate the complex interactions within the human body, including the influence of a variable human gut microbiome, genetic predispositions, and the broader dietary context29.
A significant gap in the current literature is the understanding of the “cocktail effect”. The UPFs typically contain multiple additives, and consumers are exposed to a mixture of preservatives, stabilizers, sweeteners, and emulsifiers within a single product and across their diet. Research has almost exclusively focused on the effects of single additives, yet these substances may interact synergistically or antagonistically to produce effects that are not predictable from their individual actions. The cumulative impact of this chronic, low-dose chemical cocktail on cellular metabolism remains a largely unexplored and critical area for investigation30.
To advance the field, future research should prioritize several key areas. First, there is a pressing need for more long-term human intervention studies. Randomized controlled trials comparing diets rich in UPFs to isocaloric diets comprised of minimally processed foods are crucial for isolating the effects of “processing” from confounding factors like caloric density or macronutrient composition. Such studies should incorporate sophisticated measurements of gut barrier function, systemic inflammation, multi-omics profiling of the gut microbiome, and markers of cellular metabolic health30.
Second, research must move toward investigating the synergistic effects of food additives. Advanced in vitro models, such as organ-on-a-chip systems that co-culture gut epithelial and immune cells, could offer a more physiologically relevant platform for testing the impact of additive mixtures. Third, a deeper exploration of the additive-microbiome-host axis is required to identify the specific microbial pathways that are altered by UPFs and how the resulting microbial metabolites influence host cellular processes. Finally, the development and validation of more nuanced food classification systems that may incorporate metabolic impact metrics could provide a more granular tool for both researchers and policymakers30. Addressing these research priorities will be essential for refining our understanding and developing more effective strategies to mitigate the metabolic risks posed by the global increase in UPF consumption.
CONCLUSION
The widespread integration of Ultra-Processed Foods (UPFs) into modern diets marks a significant shift in human nutrition with deep biological consequences. The industrial design of UPFs, beyond their poor nutrient profiles, actively disrupts cellular balance through additive-driven oxidative stress, mitochondrial damage, and impaired nutrient signaling. The gut microbiome plays a central role, linking dietary exposure to systemic inflammation and metabolic endotoxemia. These mechanisms collectively drive insulin resistance, fat accumulation, and energy imbalance, laying the foundation for chronic metabolic diseases. The evidence challenges traditional dietary models focused solely on nutrients and calls for a broader view that includes food processing. Public health strategies must evolve to reflect this reality by incorporating processing classifications and reassessing additive safety. Consumer education, regulatory reform, and targeted research are essential to reduce the metabolic risks posed by UPFs. Ultimately, this manuscript reframes UPFs not as passive dietary choices but as active agents of cellular disruption. Addressing their impact is critical to reversing the global rise in metabolic disease.
SIGNIFICANCE STATEMENT
Ultra-Processed Foods (UPFs) are more than nutritionally poor-hey are biologically disruptive agents that compromise cellular health. This manuscript reveals how industrial additives and processing techniques impair gut integrity, trigger systemic inflammation, and induce oxidative and mitochondrial stress. These mechanisms converge to drive insulin resistance, lipid accumulation, and metabolic dysfunction at the cellular level. By shifting the focus from nutrient content to processing impact, the research reframes UPFs as active contributors to chronic disease. It calls for a redefinition of dietary guidelines and public health strategies to address the hidden cellular toll of modern food formulations.
ACKNOWLEDGMENT
We acknowledge Federal University Wukari for providing the institutional environment that facilitated this work. We also appreciate the academic support and constructive feedback from colleagues that enhanced the manuscript.
REFERENCES
- Anastasiou, I.A., D. Kounatidis, N.G. Vallianou, A. Skourtis and K. Dimitriou et al., 2025. Beneath the surface: The emerging role of ultra-processed foods in obesity-related cancer. Curr. Oncol. Rep., 27: 390-414.
- Shahatah, F.A., T.R. Hill, A. Fairley and A.W. Watson, 2025. Ultra-processed food intakes and health outcomes in adults older than 60 years: A systematic review. Nutr. Rev., 83: 1711-1724.
- Aramburu, A., G. Alvarado-Gamarra, R. Cornejo, K. Curi-Quinto, C. del Pilar Díaz-Parra, G. Rojas-Limache and C.F. Lanata, 2024. Ultra-processed foods consumption and health-related outcomes: A systematic review of randomized controlled trials. Front. Nutr., 11.
- Singh, A. and C. Vellapandian, 2023. High risk of metabolic complications due to high consumption of processed foods. Curr. Nutr. Food Sci., 19: 198-208.
- Tompa, O., A. Kiss, S. Soós, Z. Lakner, A. Raner, G. Kasza and D. Szakos, 2025. Fifteen years of NOVA food-processing classification: “Friend or Foe” among sustainable diet indicators? A scoping review. Nutr. Rev., 83: 771-791.
- Slaathaug, C., M.M. Paulsen, S. Jafarzadeh, M.H. Carlsen and L.F. Andersen, 2024. Nutritional quality and climate impact of Norwegian adults’ diet classified according to the NOVA system. Nutr. J., 23.
- Lv, J.L., Y.F. Wei, J.N. Sun, Y.C. Shi and F.H. Liu et al., 2024. Ultra-processed food consumption and metabolic disease risk: An umbrella review of systematic reviews with meta-analyses of observational studies. Front. Nutr., 11.
- Kityo, A., B. Choi, J.E. Lee, C. Kim and S.A. Lee, 2025. Association of ultra-processed food-related metabolites with selected biochemical markers in the UK Biobank. Nutr. J., 24.
- Rondinella, D., P.C. Raoul, E. Valeriani, I. Venturini and M. Cintoni et al., 2025. The detrimental impact of ultra-processed foods on the human gut microbiome and gut barrier. Nutrients, 17.
- Atzeni, A., M.Á. Martínez, N. Babio, P. Konstanti and F.J. Tinahones et al., 2022. Association between ultra-processed food consumption and gut microbiota in senior subjects with overweight/obesity and metabolic syndrome. Front. Nutr., 9.
- Kotopoulou, S., A. Zampelas and E. Magriplis, 2023. Nitrite and nitrate intake from processed meat is associated with elevated diastolic blood pressure (DBP). Clin. Nutr., 42: 784-792.
- Martins-Gomes, C., F.M. Nunes and A.M. Silva, 2024. Natural products as dietary agents for the prevention and mitigation of oxidative damage and inflammation in the intestinal barrier. Antioxidants, 13.
- Liu, C., S. Zhan, Z. Tian, N. Li and T. Li et al., 2022. Food additives associated with gut microbiota alterations in inflammatory bowel disease: Friends or enemies? Nutrients, 14.
- Park, J.E., H.Y. Park, Y.S. Kim and M. Park, 2024. The role of diet, additives, and antibiotics in metabolic endotoxemia and chronic diseases. Metabolites, 14.
- Fadaei, M., A.S.N. Lahijan, D. Jahanmehr, A. Ahmadi and R. Asadi-Golshan, 2025. Food additives for the central nervous system, useful or harmful? An evidence-based review. Nutr. Neurosci., 28: 910-927.
- Dey, S., A. Samadder and S. Nandi, 2022. Exploring current role of nanotechnology used in food processing industry to control food additives and their biochemical mechanisms. Curr. Drug Targets, 23: 513-539.
- Jarmakiewicz-Czaja, S., A. Sokal-Dembowska and R. Filip, 2025. Effects of selected food additives on the gut microbiome and metabolic dysfunction-associated steatotic liver disease (MASLD). Medicina, 61.
- Hrncir, T., E. Trckova and L. Hrncirova, 2024. Synergistic effects of fructose and food preservatives on metabolic dysfunction-associated steatotic liver disease (MASLD): From gut microbiome alterations to hepatic gene expression. Nutrients, 16.
- Claudino, P.A., N.B. Bueno, S. Piloneto, D. Halaiko, L.P.A. de Sousa, C.H.B.J. Maia and B.D.M. Netto, 2024. Consumption of ultra-processed foods and risk for Alzheimer’s disease: A systematic review. Front. Nutr., 10.
- Harlan, T.S., R.V. Gow, A. Kornstädt, P.W. Alderson and R.H. Lustig, 2023. The metabolic matrix: Re-engineering ultraprocessed foods to feed the gut, protect the liver, and support the brain. Front. Nutr., 10.
- Galdino-Silva, M.B., K.M.M. Almeida, A.D.S. de Oliveira, J.V.L. dos Santos and M. de Lima Macena et al., 2024. A meal with ultra-processed foods leads to a faster rate of intake and to a lesser decrease in the capacity to eat when compared to a similar, matched meal without ultra-processed foods. Nutrients, 16.
- Brichacek, A.L., M. Florkowski, E. Abiona and K.M. Frank, 2024. Ultra-processed foods: A narrative review of the impact on the human gut microbiome and variations in classification methods. Nutrients, 16.
- Bassaganya-Riera, J., E.M. Berry, E.E. Blaak, B. Burlingame and J. le Coutre et al., 2021. Goals in nutrition science 2020-2025. Front. Nutr., 7.
- Abar, L., E.M. Steele, S.K. Lee, L. Kahle and S.C. Moore et al., 2025. Identification and validation of poly-metabolite scores for diets high in ultra-processed food: An observational study and post-hoc randomized controlled crossover-feeding trial. PLoS Med., 22.
- Abdelazim, A.M. and M.M. Abomughaid, 2024. Oxidative stress: An overview of past research and future insights. All Life, 17.
- Guedes, L.J.L., V.B. Tavares, S.R. Carneiro and L.M.T. Neves, 2025. The effect of physical activity on markers of oxidative and antioxidant stress in cancer patients: A systematic review and meta-analysis. BMC Cancer, 25.
- d’Alessandro, M.C.B., S. Kanaan, M. Geller, D. Praticò and J.P.L. Daher, 2025. Mitochondrial dysfunction in Alzheimer’s disease. Ageing Res. Rev., 107.
- Rong, J., Q. Yu, G. Huang, Y. Wang and N. Zhang, 2025. Advances in mitochondrial dysfunction in radiation tissue injury. Front. Physiol., 16.
- Giannakopoulos, G. and A. Prassou, 2025. Mediating and moderating mechanisms in the relationship between social media use and adolescent aggression: A scoping review of quantitative evidence. Eur. J. Invest. Health Psychol. Educ., 15.
- Nwaozuru, U., P. Murphy, A. Richard, C. Obiezu-Umeh and T. Shato et al., 2025. The sustainability of health interventions implemented in Africa: An updated systematic review on evidence and future research perspectives. Implementation Sci. Commun., 6.
How to Cite this paper?
APA-7 Style
Anih,
D.C., Tatah,
V.S., Ugwuoke,
K.C., Boyi,
R.N. (2025). Cellular Disruption by Ultra-Processed Foods: Mechanistic Pathways Linking Industrial Additives to Metabolic Disease. Research Journal of Microbiology, 20(1), 20-41. https://doi.org/10.3923/rjm.2025.20.41
ACS Style
Anih,
D.C.; Tatah,
V.S.; Ugwuoke,
K.C.; Boyi,
R.N. Cellular Disruption by Ultra-Processed Foods: Mechanistic Pathways Linking Industrial Additives to Metabolic Disease. Res. J. Microbiol 2025, 20, 20-41. https://doi.org/10.3923/rjm.2025.20.41
AMA Style
Anih
DC, Tatah
VS, Ugwuoke
KC, Boyi
RN. Cellular Disruption by Ultra-Processed Foods: Mechanistic Pathways Linking Industrial Additives to Metabolic Disease. Research Journal of Microbiology. 2025; 20(1): 20-41. https://doi.org/10.3923/rjm.2025.20.41
Chicago/Turabian Style
Anih, David, Chinonso, Verwiyeh Silas Tatah, Kenneth Chinekwu Ugwuoke, and Richard-Harris Nsenreuti Boyi.
2025. "Cellular Disruption by Ultra-Processed Foods: Mechanistic Pathways Linking Industrial Additives to Metabolic Disease" Research Journal of Microbiology 20, no. 1: 20-41. https://doi.org/10.3923/rjm.2025.20.41

This work is licensed under a Creative Commons Attribution 4.0 International License.